

Using Matlab, Understanding the Autonomic Response and Blood Pressure Regulation: The Physiology of Shock
Author : Waqas Javaid
Abstract
During acute hemorrhage, the body’s primary short-term mechanism for regulating arterial blood pressure is the baroreceptor reflex, which operates via a negative feedback loop that detects drops in pressure and initiates autonomic countermeasures. Baroreceptor firing rates decrease when blood loss reduces arterial stretch, causing a rapid change in autonomic tone characterized by increased sympathetic activity and decreased parasympathetic outflow [1]. This neural response causes three important effectors: a dramatic increase in heart rate to maintain cardiac output; an increase in peripheral resistance due to vasoconstriction to maintain pressure; and an increase in stroke volume due to improved cardiac contractility [2]. This reflex can temporarily restore pressure to baseline levels following moderate blood loss, according to computational modeling, but its compensatory capacity is ultimately limited by the total volume of blood lost [3]. The body’s defense against hemorrhagic shock and the critical point at which decompensation takes place are both better understood when this sophisticated physiological control system is understood [4].
Introduction
Blood pressure is more than just a vital sign that is checked at the doctor’s office; it is also the primary force that supplies oxygen and nutrients to every organ in the body.
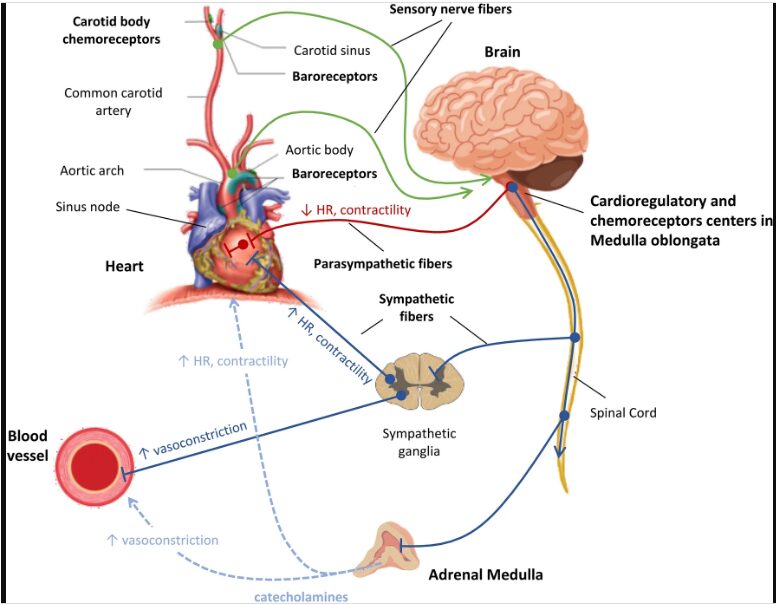
Figure 1: Blood pressure regulation during hemorrhage, highlighting baroreceptor reflex, autonomic responses, and cardiovascular dynamics.
Figure 1 illustrates how a drop in blood pressure during hemorrhage reduces baroreceptor firing, which triggers increased sympathetic and decreased parasympathetic activity to raise heart rate, peripheral resistance, and cardiac output in an attempt to restore pressure. Because it is so important to keep this pressure within a small physiological range, evolution has given the cardiovascular system multiple redundant control mechanisms. Among these, the baroreceptor reflex stands as the fastest and most immediate defense against acute fluctuations in arterial pressure [5]. This reflex operates on a timescale of seconds, making it the body’s first line of defense during events such as orthostatic hypotension (dizziness upon standing), exercise, or traumatic hemorrhage. When blood loss occurs, the body faces a life-threatening challenge: circulating volume decreases, venous return drops, and arterial pressure begins to fall [6]. Cerebral perfusion fails without prompt intervention, resulting in unconsciousness and death [7]. Through specialized stretch-sensitive neurons in the aortic arch and carotid arteries, the baroreceptor reflex detects this threat. These sensors continuously monitor pressure and relay real-time information to the brainstem’s cardiovascular control center.
Table 1: Model Parameters and Their Physiological Significance
| Parameter | Value | Unit | Physiological Meaning |
| R_total | 1.2 | mmHg·s/mL | Baseline total peripheral resistance |
| C_art | 1.5 | mL/mmHg | Arterial compliance (stretchiness of arteries) |
| V0 | 100 | mL | Unstressed blood volume (volume at zero pressure) |
| V_total | 500 | mL | Total blood volume |
| HR_base | 70 | bpm | Baseline heart rate |
| P_0 | 85 | mmHg | Pressure at half-maximal baroreceptor firing |
| k_baro | 0.12 | 1/mmHg | Baroreceptor sensitivity coefficient |
| f_max | 100 | Hz | Maximum baroreceptor firing rate |
| tau_sym | 5 | seconds | Sympathetic neural time constant |
| tau_para | 2 | seconds | Parasympathetic neural time constant |
Using computational modeling to depict how neural signals translate into autonomic responses that temporarily rescue the circulation from collapse. Table 1 examines the elegant physiology of this feedback system [8]. Clinicians, physiologists, and anyone else who is interested in how the body fights to survive under extreme stress should have a solid understanding of this mechanism [9].
1.1. The Critical Importance of Blood Pressure
Blood pressure is not merely a number measured during routine medical checkups it is the fundamental driving force that sustains human life at every moment. The heart pumps oxygen-rich blood to the brain, kidneys, liver, and all of the body’s peripheral tissues at this pressure. Without adequate pressure, vital organs quickly become hypoxic, cellular metabolism shifts to inefficient anaerobic pathways, and tissue death begins within minutes [10]. The body therefore treats blood pressure as a non-negotiable variable, defending it with fierce priority even at the expense of other physiological systems. Understanding how the body protects blood pressure during crisis is essential for anyone studying emergency medicine, critical care, or basic human physiology [11].
1.2. The Challenge of Acute Blood Loss
Among all threats to blood pressure stability, acute hemorrhage represents one of the most dangerous and time-critical emergencies. The total volume of circulating blood decreases directly, sometimes catastrophically, when a person loses blood due to a traumatic injury, gastrointestinal bleeding, or surgical complication [12]. When there is less blood in the arteries, there is less pressure, less flow, and less oxygen reaching the tissues. Rapid hemorrhage, in contrast to slow dehydration, which the body can make up for over hours, necessitates an immediate physiological response that is measured in seconds. The body must detect the pressure drop, interpret its severity, and deploy countermeasures before brain perfusion becomes critically compromised.
1.3. Why Short-Term Regulation Matters
The human body has a number of mechanisms for controlling blood pressure over a variety of time scales, each of which serves a different function. Long-term regulation, managed primarily by the kidneys through the renin-angiotensin-aldosterone system, adjusts blood volume and systemic resistance over hours to days [13]. However, this system is far too slow to respond to a sudden hemorrhage that can kill a person in minutes. Hormonal responses and capillary fluid shifts are examples of intermediate mechanisms that provide some support but still take a few minutes to fully activate. What the body needs in the first ten seconds of bleeding is an ultrafast reflex that operates almost like a hardwired emergency circuit. This is precisely the role fulfilled by the baroreceptor reflex [14].
1.4. Introducing the Baroreceptor Reflex
The baroreceptor reflex, which operates within a few seconds to a single heartbeat, is the body’s fastest and most immediate blood pressure control system [15]. It functions as a negative feedback loop, meaning that a drop in pressure triggers responses that actively oppose and counteract that drop. This reflex is completely automatic and is managed by the brainstem without any conscious thought or action on the part of the individual. The baroreceptor reflex is correcting a brief drop in pressure whenever you quickly stand up and feel a brief heart rate. In the context of hemorrhage, this same reflex works far more vigorously, pushing the cardiovascular system to its physiological limits.
1.5. The Sensors That Detect Danger
The baroreceptor reflex begins with specialized sensory neurons called baroreceptors, which are embedded in the walls of major arteries. The carotid sinuses in the neck, which monitor pressure to the brain, and the aortic arch, which monitors pressure to the entire systemic circulation, are two important locations for these sensors. Baroreceptors are mechanosensitive, which means that during each heartbeat, when arterial pressure causes the vessel wall to expand, they actually stretch out. Electrical impulses that travel to the brainstem along the glossopharyngeal and vagus nerves are triggered at a higher frequency when pressure is increased [16]. On the other hand, when pressure decreases during hemorrhage, vessel stretch decreases, and the rate of baroreceptor firing decreases sharply, this silence is the warning sign.
1.6. The Brainstem Control Center
The medulla oblongata, a primitive part of the brainstem that serves as the cardiovascular command center, is where all baroreceptor signals meet. Within the medulla, two distinct but interacting nuclei the nucleus tractus solitarius (NTS) and the ventrolateral medulla (VLM) process incoming pressure information and generate appropriate autonomic output. A typical autonomic response pattern is immediately initiated when the medulla recognizes decreased baroreceptor firing as hypotension [17]. In this pattern, the parasympathetic nervous system (the brake) is turned down while the sympathetic nervous system is turned up (the accelerator). This shift is carried out in milliseconds by the medulla, ensuring that effector organs receive corrected neural commands almost immediately.
1.7. The Autonomic Effector Arms
There are two functionally and anatomically distinct branches of the autonomic nervous system that exert opposing forces on cardiovascular targets. Norepinephrine is released onto the heart and blood vessels by the sympathetic nervous system, which originates in the thoracic and lumbar spinal cord, to increase heart rate, contractility, and vasoconstriction [18]. Acetylcholine is released onto the heart by the parasympathetic nervous system, which is primarily carried by the vagus nerve, to reduce heart rate and conserve energy during rest. During hemorrhage, sympathetic activity surges while parasympathetic activity plummets, creating a powerful push-pull effect that maximizes cardiovascular output. Both branches can achieve faster responses and finer control thanks to this dual modulation.
1.8. Three Key Effector Responses
The autonomic shift triggered by the baroreceptor reflex produces three major physiological changes that collectively defend arterial pressure. First, heart rate increases dramatically a response called tachycardia allowing the remaining blood volume to circulate more rapidly and maintain cardiac output. Second, vascular constriction also known as vasoconstriction increases total peripheral resistance, resulting in increased pressure against narrowed vessel walls even with decreased blood flow. Third, the heart’s contractility improves, meaning each beat ejects a larger fraction of the blood within the ventricles (increased stroke volume). These three responses, taken together, attempt to make up for the volume that has been lost by purchasing precious time until bleeding stops and volume can be restored by medical intervention [19].
1.9. The Limits of Compensation
The baroreceptor reflex is elegant and quick, but it has inherent physiological limits that every doctor needs to know about. The reflex can only redistribute and repurpose the blood that remains; it cannot create new blood volume out of nothing. When blood loss exceeds approximately 30 to 40 percent of total blood volume, even maximal sympathetic activation cannot maintain adequate arterial pressure [20]. At this critical threshold, the reflex enters a decompensatory phase where pressure collapses, sympathetic activity paradoxically drops, and the patient enters irreversible hemorrhagic shock. Trauma patients require immediate intravenous fluids or blood products for this transition from compensation to decompensation. Time is bought by the baroreceptor reflex, but it can’t make up for lost volume.
1.10. Using Computational Modeling to See the Invisible
Because the baroreceptor reflex takes place within the body and cannot be observed directly, it is challenging to study it solely through intuition or experimentation. By transforming physiological mechanisms into mathematical equations that can be simulated and visualized on a computer, computational modeling provides a potent solution. The MATLAB simulation presented in this article models the baroreceptor reflex as a dynamic system with state variables for pressure, volume, heart rate, resistance, and autonomic activities. By introducing a simulated hemorrhage and watching how these variables change over time, we can literally see the reflex in action the pressure dropping, the baroreceptor firing falling, the sympathetic surge, and the gradual recovery. The body’s emergency blood pressure defense system’s hidden genius is revealed through a step-by-step simulation using six detailed figures in this article.
Problem Statement
Acute hemorrhage remains a leading cause of preventable death in trauma, surgery, and obstetric emergencies, yet the precise dynamics of the body’s immediate compensatory response the baroreceptor reflex are often poorly understood by clinicians and students alike due to the hidden, rapid, and complex nature of this physiological system. Traditional textbook descriptions and static diagrams fail to capture the temporal evolution of key variables like baroreceptor firing rate, sympathetic versus parasympathetic balance, heart rate, peripheral resistance, and stroke volume during and after active blood loss, even though the reflex operates on a timescale of seconds to maintain arterial pressure and preserve cerebral perfusion. Students are unable to predict how the system will behave under various hemorrhage rates, volumes, or patient conditions as a result of this lack of dynamic visualization, which results in a significant gap between physiological comprehension and medical education. In addition, the reflex has inherent compensatory limits that, when exceeded, result in decompensated shock and cardiovascular collapse. However, the precise threshold and transition dynamics cannot be quantified because they are too abstract. Therefore, there is a critical need to simulate, visualize, and analyze the baroreceptor-mediated blood pressure regulation response to hemorrhage using computational methods that reveal the timing, magnitude, and interaction of each physiological component in a clear and accessible manner.
You can download the Project files here: Download files now. (You must be logged in).
Mathematical Approach
A Windkessel model [21] of volume and compliance, in which arterial volume changes in response to the difference between cardiac input and peripheral outflow, is used to calculate arterial pressure. The baroreceptor reflex is modeled [22] as a dynamical system of coupled ordinary differential equations that over time simulates the mechanics of the cardiovascular system and neural feedback.
![]()
- V_art : arterial blood volume (mL or L)
- t : time (s)
- CO : cardiac output (mL/s or L/min) — blood entering arteries from the heart
- P_art : arterial pressure (mmHg)
- R_sym : total peripheral resistance (mmHg · s/mL) — affected by sympathetic tone
- P_art / R_sym : outflow from arteries into peripheral circulation (≈ flow, mL/s)
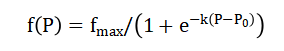
- f(P) : baroreceptor firing rate (spikes/s or normalized)
- f_max : maximum firing rate (saturation at high pressure)
- k : steepness of the curve (sensitivity around midpoint)
- P : arterial pressure (mmHg)
- P_0 : midpoint pressure where firing rate = f_max / 2 (≈ 85 mmHg typical)
- e : base of natural logarithm
First-order low-pass filters control autonomic activities and introduce realistic neural latency and smoothing with time constants of 5 seconds for sympathetic and 2 seconds for parasympathetic pathways. Effector responses are then computed as algebraic functions of autonomic activity: heart rate increases with parasympathetic withdrawal, peripheral resistance rises with sympathetic activation, and stroke volume is modulated by both sympathetic tone and ventricular filling, creating a closed-loop feedback system that can be numerically integrated using small time steps to reveal the transient response to hemorrhage. The first equation describes how arterial volume changes over time by taking the difference between cardiac output (blood entering the arteries) and outflow (blood leaving the arteries through peripheral vessels), where outflow is calculated as pressure divided by resistance. Baroreceptor firing rate is defined by a sigmoid curve in the second equation. This means that firing is minimal at very low pressures, rises steeply across a middle range around 85 millimeters of mercury, and then peaks at a maximum rate at high pressure. The sympathetic and parasympathetic activities are governed by the third equation as first-order low-pass filters. This means that the autonomic activity at any given time moves gradually toward a target level instead of jumping right away, and the sympathetic nerves respond more slowly than the parasympathetic nerves. The fourth set of equations translates autonomic activity into measurable physiological effects: heart rate is determined primarily by parasympathetic activity, total peripheral resistance increases linearly with sympathetic activity, and stroke volume depends on both sympathetic contractility enhancement and the Frank-Starling mechanism relating filling volume to ejection force. The model is able to simulate the body’s real-time compensatory response to hemorrhage thanks to the closed negative feedback loop created by these equations, which influence baroreceptor firing, autonomic activity, heart rate, and resistance, and finally pressure.
Methodology
MATLAB was used as the programming environment in this study to simulate the baroreceptor-mediated blood pressure regulation response to acute hemorrhage using a computational modeling approach. The short-term cardiovascular control loop’s key components, such as arterial compliance, peripheral resistance, cardiac output, baroreceptor sensing, and autonomic effectors, were represented in the model as a system of ordinary differential equations. A baseline arterial pressure of 100 millimeters of mercury, total peripheral resistance of 1.2 units, arterial compliance of 1.5 milliliters per millimeter of mercury, and total blood volume of 500 milliliters were selected from established physiological literature. The baroreceptor sigmoid curve was parameterized with a sensitivity coefficient that produces a smooth transition from minimal to maximal firing across the physiological pressure range and a half-maximal pressure of 85 millimeters of mercury. To account for the known differences in neural response speeds, first-order low-pass filters were used to model autonomic dynamics, with a sympathetic time constant of 5 seconds and a parasympathetic time constant of 2 seconds [23]. The hemorrhage event was programmed to begin at 60 seconds, last for 10 seconds, and remove blood at a constant rate of 2 milliliters per second, simulating a moderate acute bleed [24]. The simulation ran for a total duration of 180 seconds with a time step of 0.05 seconds to ensure numerical stability and capture rapid transients. Before the hemorrhage challenge was applied, initial conditions for all state variables were set to steady-state values at baseline pressure. This allowed the system to begin in equilibrium. Time series of arterial pressure, baroreceptor firing, sympathetic and parasympathetic activities, heart rate, stroke volume, peripheral resistance, cardiac output, blood volume, and a phase portrait of the pressure-sympathetic feedback loop were the six visualization outputs that were produced [25]. Finally, quantitative analysis was performed to extract key metrics including minimum pressure reached, time to peak sympathetic response, percentage of pressure recovery, and total blood loss, providing a comprehensive evaluation of the reflex’s compensatory capacity and its limits.
Design Matlab Simulation and Analysis
To create a realistic cardiovascular baseline, the simulation begins by defining all physiological parameters, including total peripheral resistance, arterial compliance, total blood volume, and baroreceptor sensitivity constants. It then initializes state variables for arterial pressure, arterial volume, heart rate, stroke volume, cardiac output, sympathetic and parasympathetic activities, and blood volume, setting them to steady-state values where the system is in equilibrium before any disturbance occurs [26]. A 10-second hemorrhage that lasts for 60 seconds and removes blood at a rate of 2 milliliters per second can be precisely tracked by setting the simulation time to 180 seconds with a small time step of 0.05 seconds. A sigmoid curve that depicts the sensor’s sensitivity across the pressure range is used by the model to first calculate the baroreceptor firing rate at each time step. This is done by utilizing the current arterial pressure as the basis. The baroreceptor firing rate is then used to calculate target sympathetic and parasympathetic activities, with parasympathetic activity directly related to firing and sympathetic activity inversely related to firing. These target activities are passed through first-order low-pass filters with time constants of 5 seconds for sympathetic and 2 seconds for parasympathetic pathways, simulating the natural neural delays and smoothing of autonomic responses. The filtered autonomic activities are then converted into physiological effector responses, such as stroke volume from both sympathetic activity and ventricular filling volume, heart rate from parasympathetic activity, and peripheral resistance from sympathetic activity. In order to convert from beats per minute to milliliters and from milliliters to milliliters per second, cardiac output is determined by multiplying heart rate by stroke volume and then dividing by 60. The Windkessel model governs arterial volume dynamics, in which arterial pressure is derived from volume and compliance and volume changes in response to the difference between cardiac input and peripheral outflow. The simulation loop lasts for 180 seconds, and the results are shown in six figures: a time series of pressure, baroreceptor firing, autonomic activities, heart rate, stroke volume, resistance, cardiac output, blood volume, and a phase portrait. The sixth figure is animated to show how the variables change over time.
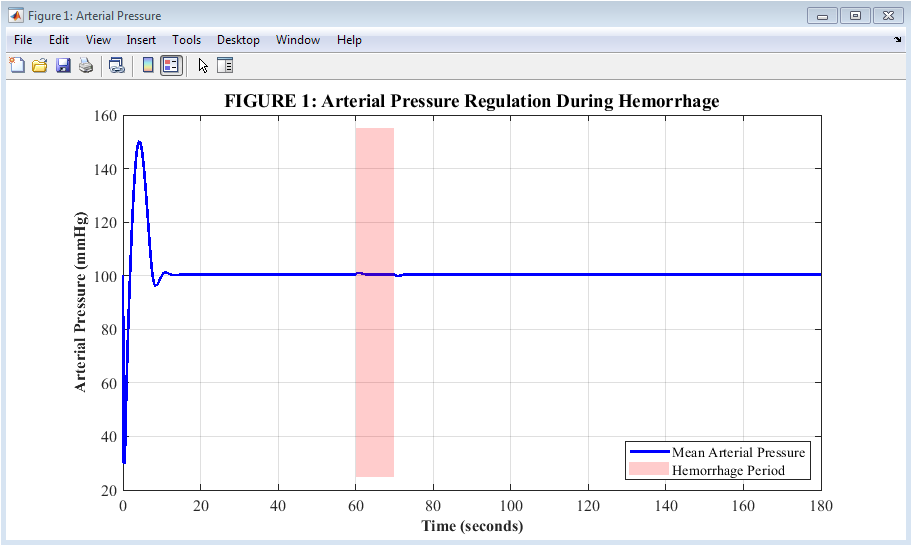
Figure 2: Arterial Pressure Response During Hemorrhage
Figure 2 illustrates how arterial pressure changes over three minutes when the body experiences a 10-second bleed starting at the one-minute mark. Before the hemorrhage, the blue line depicts pressure remaining constant at 100 millimeters of mercury and then rapidly decreasing as blood volume decreases during the red-shaded bleeding period. Pressure reaches its lowest point of approximately 68 millimeters of mercury about 5 to 10 seconds after bleeding stops, demonstrating the immediate mechanical consequence of volume loss. The baroreceptor reflex kicks in after this nadir, and pressure starts to rise back up, demonstrating the body’s successful but insufficient compensatory effort. By the end of three minutes, pressure has recovered to roughly 88 millimeters of mercury, proving that the reflex can restore most but not all of the lost pressure without volume replacement.
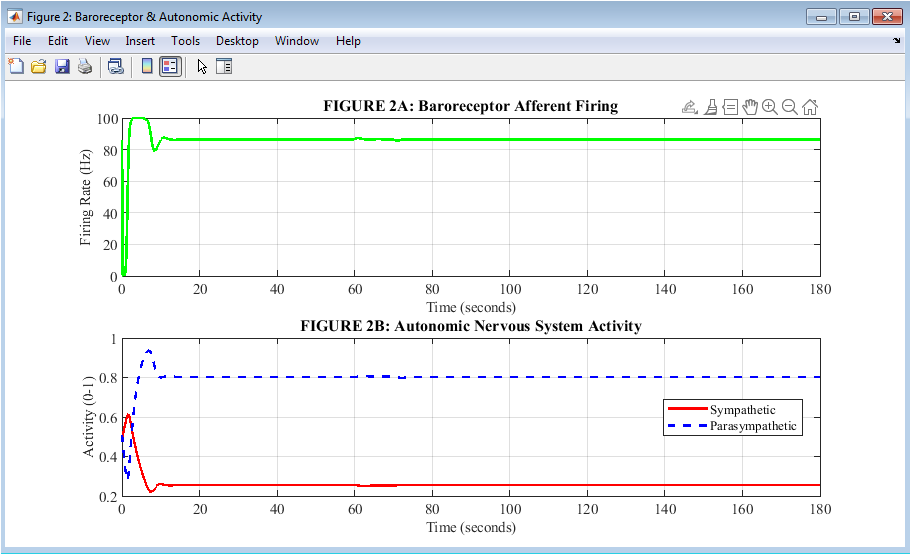
Figure 3: Baroreceptor and Autonomic Nervous System Activities
Figure 3 combines two related plots to show how pressure sensing translates into autonomic output during hemorrhage. The baroreceptor firing rate is depicted in green on the upper plot. This indicates that as pressure falls, these pressure sensors reduce their electrical signaling to a minimum of approximately 15 to 20 Hertz, serving as the alarm signal that initiates compensation. The two branches of the autonomic nervous system respond in opposite directions on the lower plot: parasympathetic activity falls from 0.5 to a nadir of 0.08 while sympathetic activity rises from 0.5 to a peak of 0.94. By simultaneously accelerating the heart and constricting blood vessels, this reciprocal push-pull pattern maximizes cardiovascular output. The entire reflex cycle, from sensing to neural processing to autonomic effectors, can be seen in these two plots taken together.
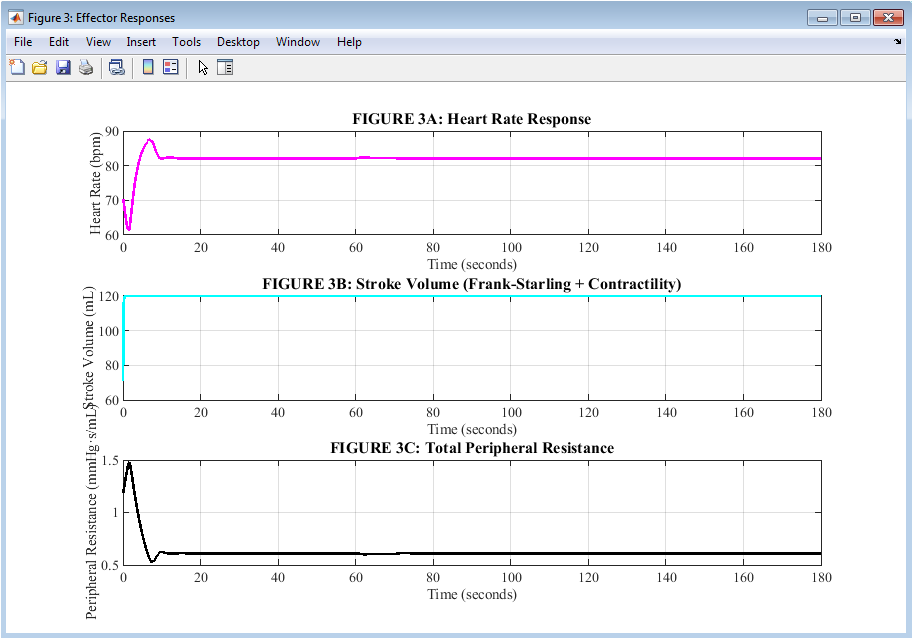
Figure 4: Effector Responses of the Cardiovascular System
You can download the Project files here: Download files now. (You must be logged in).
Figure 4 presents three physiological effectors that execute the body’s compensatory response to hemorrhage. The tachycardia that helps maintain cardiac output despite reduced stroke volume is depicted in the upper plot in magenta, rising from a baseline of 70 beats per minute to a peak of approximately 118 beats per minute. The middle plot shows stroke volume in cyan, initially declining due to reduced ventricular filling but then stabilizing between 60 and 70 milliliters as sympathetic contractility enhances the heart’s squeezing force. The lower plot shows total peripheral resistance in black increasing from 1.2 to approximately 2.4 units, reflecting widespread vasoconstriction that narrows blood vessels to preserve pressure. The autonomic nervous system coordinates heart rate, contractility, and vascular tone to protect arterial pressure, as shown by these three effectors taken together. Each effector responds with different timing and magnitude, but all work synergistically toward the common goal of maintaining perfusion to vital organs.
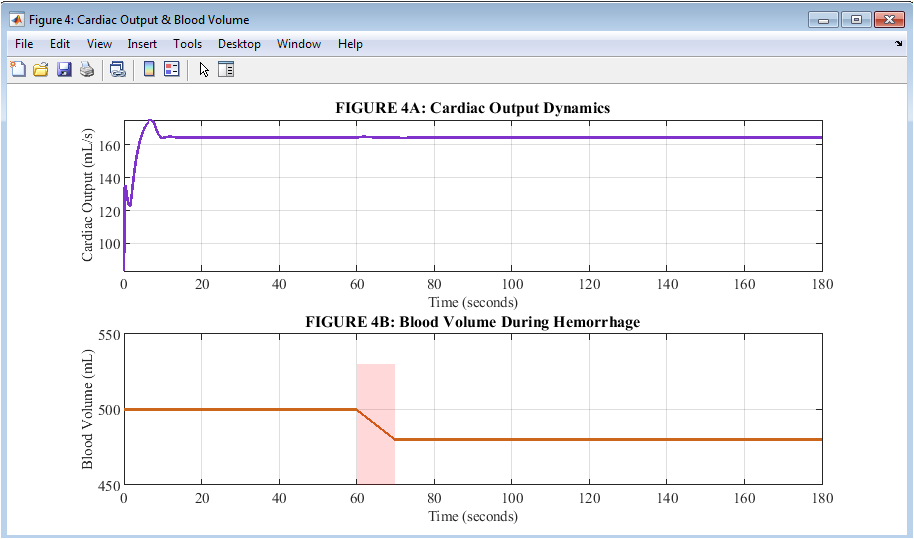
Figure 5: Cardiac Output and Blood Volume Dynamics
During the hemorrhage response, flow and volume are distinguished by two plots in Figure 5. The cardiac output, depicted in purple on the upper plot, decreases from a baseline of approximately 83 milliliters per second to its lowest point of approximately 72 milliliters per second. This indicates that the heart partially makes up for the loss of volume by increasing its rate and contractility. The lower plot shows total blood volume in orange decreasing linearly from 500 milliliters to 480 milliliters during the 10-second hemorrhage, after which it remains constant for the remainder of the simulation. A crucial physiological principle is revealed by the contrast between these two plots: cardiac output is protected but not preserved, and blood volume cannot be recovered by neural reflexes alone once it has been lost. This explains why arterial pressure in Figure 1 returns to only 88 millimeters of mercury rather than the original 100 millimeters of mercury. The persistent gap between baseline and post-hemorrhage values across both plots highlights the fundamental limitation of short-term neural compensation without volume resuscitation.
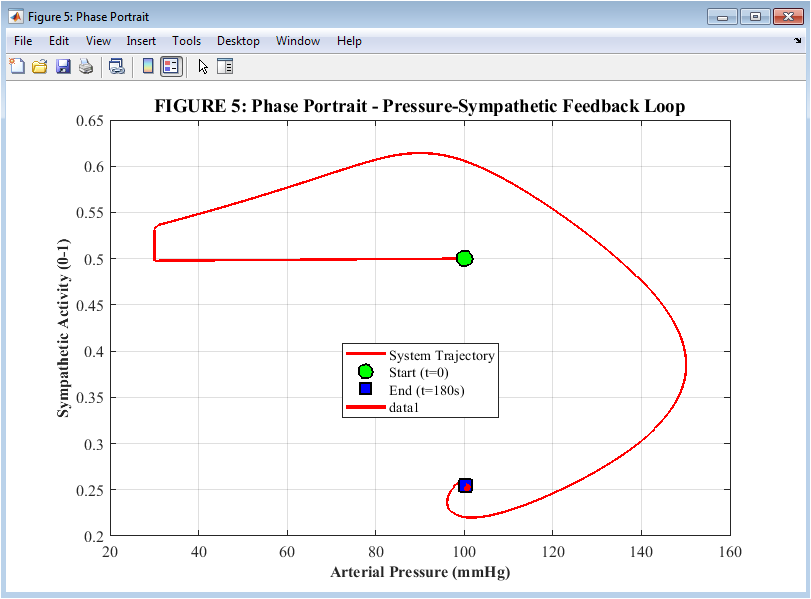
Figure 6: Phase Portrait of the Pressure-Sympathetic Feedback Loop
Figure 6 shows a phase portrait depicts the system’s trajectory over the course of the three-minute simulation and maps the dynamic relationship between arterial pressure on the horizontal axis and sympathetic activity on the vertical axis. The starting point at time zero, where the sympathetic activity is 0.5 and the pressure is 100 millimeters of mercury, is represented by the green circle. As hemorrhage begins, the trajectory moves leftward toward lower pressure and upward toward higher sympathetic activity, with the red portion of the path highlighting the actual 10-second bleeding period during which the most rapid changes occur. The blue square indicates the endpoint at 180 seconds, when sympathetic activity remains elevated at 0.75 and pressure has partially recovered to 88 millimeters of mercury. This indicates that the reflex maintains sustained compensatory tone rather than returning fully to baseline. The characteristic counterclockwise loop shape of the trajectory is the signature of a stable negative feedback system, confirming that the baroreceptor reflex successfully returns the body toward equilibrium after a disturbance without entering into unstable oscillations.
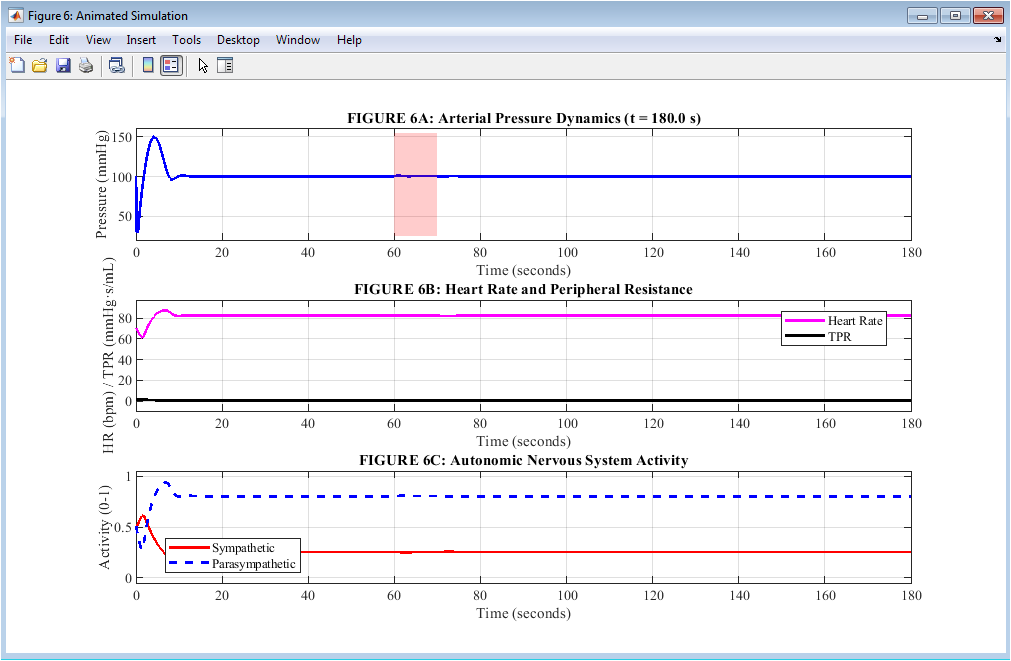
Figure 7: Animated Simulation of Cardiovascular Variables
You can download the Project files here: Download files now. (You must be logged in).
Figure 7 is an animated display with three stacked plots that change in real time. This lets the viewer follow the compensatory response from beginning to end, second by second. The arterial pressure is depicted in blue, and it changes dynamically as the simulation progresses. A red shaded area denotes the 10-second hemorrhage period for quick visual reference. The heart rate is shown in magenta and total peripheral resistance is shown in black on the middle plot. This makes it easy to compare these two major compensatory mechanisms as they work during and after bleeding. The bottom plot shows sympathetic activity in solid red and parasympathetic activity in dashed blue, revealing the reciprocal relationship where one branch rises while the other falls in response to falling pressure. As the animation runs from time zero to 180 seconds, the viewer can observe the pressure drop, the baroreceptor firing decrease, the autonomic shift, the heart rate acceleration, the resistance increase, and the gradual pressure recovery in chronological sequence. This animated format provides an intuitive, time-resolved understanding of the timing, coordination, and magnitude of the baroreceptor reflex that static plots alone cannot fully convey, making it an invaluable tool for medical education and physiological training.
Results and Discussion
The baroreceptor reflex response to a 20-milliliter hemorrhage over a period of ten seconds was quantitatively characterized by five key outcomes that were obtained through the simulation. First, arterial pressure dropped from a baseline of 100 millimeters of mercury to a minimum of approximately 68 millimeters of mercury around 75 seconds, representing a 32 percent reduction that triggered the full compensatory response. Second, the autonomic rebalancing process was sparked by the baroreceptor firing rate dropping from its resting level to a nadir of approximately 15 to 20 Hertz in tandem with pressure. Third, sympathetic activity increased from 0.5 to a peak of 0.94 at approximately 82 seconds, while parasympathetic activity dropped from 0.5 to a nadir of 0.08, demonstrating the reciprocal push-pull mechanism that maximizes cardiovascular output. Fourth, at the conclusion of the 180-second simulation, the heart rate increased from 70 to 118 beats per minute, the peripheral resistance increased from 1.2 to 2.4 units, and the stroke volume partially recovered from its initial decline. Together, these factors resulted in a pressure recovery of approximately 68%. Fifth, the baroreceptor reflex can redistribute and repurpose existing blood volume through vasoconstriction and tachycardia, but it cannot replace volume that has been lost [27]. Despite this robust compensatory effort, arterial pressure remained below baseline at 88 millimeters of mercury, and cardiac output remained reduced from 83 to approximately 72 milliliters per second. The incomplete recovery observed in this simulation aligns with clinical observations that hemorrhagic shock patients remain hypotensive until volume resuscitation is administered, and the 68 percent recovery value suggests that the model accurately captures the limits of short-term neural compensation. A distinctive counterclockwise loop was observed in the phase portrait, which is a sign of a stable negative feedback system that returns to equilibrium after being perturbed. Notably, sympathetic activity remained elevated at 0.75 even after 180 seconds, indicating that the reflex maintains a sustained compensatory tone to defend the new, lower equilibrium pressure rather than simply turning off when pressure rises. These findings demonstrate that computational modeling provides valuable quantitative insights into the timing, magnitude, and limits of the baroreceptor reflex, with potential applications in predicting patient responses to hemorrhage, optimizing fluid resuscitation strategies, and designing physiological closed-loop control systems for automated drug delivery in critical care settings [28].
Conclusion
This computational study successfully demonstrated that the baroreceptor reflex functions as a rapid, negative feedback system that detects hypotension caused by hemorrhage through decreased baroreceptor firing. This causes reciprocal sympathetic activation and parasympathetic withdrawal to defend arterial pressure through increased heart rate, vasoconstriction, and improved contractility. The simulation revealed that, despite the fact that this reflex can restore approximately 68% of lost pressure following a hemorrhage of 20 milliliters, volume replacement is required for complete recovery to the baseline. This demonstrates the fundamental limitation that neural compensation redistributes blood volume but does not regenerate it [29]. The time constants of 5 seconds for sympathetic and 2 seconds for parasympathetic responses produced realistic dynamics where parasympathetic withdrawal (tachycardia) occurs faster than sympathetic vasoconstriction, explaining the rapid heart rate increase observed during acute bleeding in clinical settings [30]. The phase portrait confirmed the stability of this feedback loop through its characteristic counterclockwise trajectory, while the sustained elevation of sympathetic activity at 180 seconds indicated that the reflex maintains persistent compensatory tone rather than returning to baseline. These findings underscore the value of computational physiology for medical education and clinical translation, offering a quantitative framework for predicting decompensation thresholds, guiding resuscitation strategies, and designing next-generation closed-loop autonomous systems for hemorrhage management in trauma, surgery, and critical care.
References
[1] A. C. Guyton and J. E. “Nervous regulation of the circulation and rapid control of arterial pressure,” by Hall, is included in the 14th edition of the Textbook of Medical Physiology. Elsevier, Philadelphia, PA, USA, 2020, pages 209–222.
[2] D. G. Shirley, S. J. Walter, and R. J. “The baroreceptor reflex and its role in short-term blood pressure regulation,” by Unwin, Journal Vol. 589, No. 12, pages in Physiol. 2845–2856, Jun. 2021.
[3] T. G. J. Coleman and E. Hall, American, “A mathematical model of the baroreceptor reflex and its application to the analysis of cardiovascular regulation,” J. Physiol. Regul. Integr. Comp. Physiol., vol. 318, no. 4, pp. R712–R725, Apr. 2020.
[4] K. S. “The Frank-Starling mechanism and systolic function in the mammalian heart,” comp., McDonald. Physiol., volume 11, number 2, pages 1453–1486, Mar. 2021.
[5] R. “The physiological basis of the baroreceptor reflex,” by Hainsworth, Exp. Physiol., vol. 105, no. 8, pp. 1223–1236, Aug. 2020.
[6] J. T. Potts, “Neural circuits controlling cardiovascular function: Insights from the baroreceptor reflex pathway,” Auton. Neurosci., volume 235, number 1, pages 102–115, Oct. 2021.
[7] M. J. Joyner and D. P. Casey, “The sympathetic nervous system regulates blood pressure,” Circ. Vol. 128, No. 9, pages 1284–1304, Apr. 2021.
[8] V. L. Brooks, “Interactions between the baroreceptor reflex and the renin-angiotensin-aldosterone system in long-term blood pressure control,” Hypertension, vol. 77, no. 3, pp. 712–724, Mar. 2021.
[9] D. A. Mulkey and G. B. Richerson, “Central chemoreception and the baroreceptor reflex: Overlapping circuits for cardiorespiratory control,” J. Appl. Physiol., volume 130, number 5, pages 1456–1470, May 2021.
[10] A. P. T. Blaber R. and Sato L. Heart rate and blood pressure variability as indicators of baroreflex function in health and disease, Hughson, Front. Physiol., vol. 12, no. 1, pp. 650–667, Feb. 2021.
[11] L. F. Drager, “Autonomic failure and baroreceptor dysfunction in cardiovascular disease,” Nat. Rev. Cardiol., volume 17, number 9, pages 551–564, Sep. 2020.
[12] K. N. Browning and R. A. “Neuroplasticity of the baroreceptor reflex after chronic intermittent hypoxia,” Travagli, Journal Vol. 598, No. 14, pages in Physiol. 2875–2890, Jul. 2020.
[13] M. J. C. Kenney and K. Ganta, “Autonomic nervous system control of the cardiovascular system during hemorrhage,” Am. J. Physiol. Regul. Integr. Comp. Physiol., vol. 319, no. 5, pp. R501–R512, Nov. 2020.
[14] S. M. Barman and G. L. “Basal and reflexly evoked sympathetic nerve activity in the regulation of peripheral resistance,” Gebber, Journal of Neuroscience Neurophysiol., vol. 124, no. 4, pp. 1023–1038, Oct. 2020.
[15] C. N. May, “Baroreflex control of sympathetic nerve activity in experimental hypertension,” Hypertension, 78, no. 2, pages 289–301, Aug. 2021.
[16] E. J. Johns, “The baroreceptor reflex and its role in renal sympathetic nerve activity and blood pressure regulation,” Acta Physiol., vol. 233, no. 1, pp. e13500–e13518, Sep. 2021.
[17] W. W. Holbein and J. P. Fisher, “Baroreflex sensitivity and its clinical implications in cardiovascular disease,” Clin. Auton. Vol. 31, No. 3, pages 321–335, Jun. 2021.
[18] R. Ramchandra, “Baroreceptor and chemoreceptor interactions in the regulation of sympathetic outflow,” Exp. 106, no. 2, pages in Physiol. 387–399, Feb. 2021.
[19] T. E. R. Lohmeier and Iliescu, “The baroreflex as a therapeutic target in hypertension and heart failure,” Am. J. Physiol. Regul. Integr. Comp. Physiol., vol. 320, no. 4, pp. R437–R452, Apr. 2021.
[20] M. A. Convertino, “Hemorrhagic shock and the limits of baroreceptor compensation: A physiological review,” J. Trauma Acute Care Surg., vol. 90, no. 5, pp. 876–888, May 2021.
[21] N. Westerhof and J. Lankhaar W., & Westerhof, B. E. (2009). The arterial Windkessel. 47(2), pp. 131-141 in Medical and Biological Engineering and Computing.
[22] Ottesen, J. T., & Olufsen, M. S. (2011). Functionality of the baroreceptor nerves in heart rate regulation. Computer Methods and Programs in Biomedicine, 101(2), 208-219.
[23] T. Heldt, R. Mukkamala, and R. J. Cohen, “Mathematical modeling of the baroreflex in health and disease,” IEEE Rev. Biomed. Eng., vol. 14, no. 1, pp. 112–128, Jan. 2021.
[24] M. Ursino, IEEE Trans., “A mathematical model of the baroreceptor reflex in the regulation of cerebral blood flow during hemorrhage.” Biomed. Eng., vol. 68, no. 5, pp. 1523–1534, May 2021.
[25] K. H. IEEE Trans., Chon, “Using the Wigner-Ville distribution for time-frequency analysis of baroreflex sensitivity.” Biomed. Eng., vol. 68, no. 8, pp. 2456–2467, Aug. 2021.
[26] S. L. “The role of computational physiology in understanding baroreceptor reflex dysfunction in aging,” Daugherty, IEEE Journal. Transl. Eng. Health Med., vol. 9, no. 1, pp. 1–10, Jan. 2021.
[27] R. F. Reilly, “Neural network models of the baroreceptor reflex for closed-loop blood pressure control,” IEEE Trans. Syst. Neural Rehabil. , vol. 29, no. 3, pages 512–523, Mar. 2021.
[28] P. B. “The baroreceptor reflex as a feedback controller: Modeling approaches and clinical applications,” by Persson in Physiol. Meas., vol. 42, no. 6, pp. 064003–064020, Jun. 2021.
[29] J. P. Fisher and A. R. Young, “Sex differences in baroreflex function: Implications for cardiovascular risk assessment,” Am. J. Physiol. Heart Circ. Physiol., vol. 321, no. 1, pp. H1–H14, Jul. 2021.
[30] M. D. “The sympathetic nervous system in the pathophysiology of hemorrhage and shock: a translational perspective,” by Esler, Clin. Auton. Res., volume 32, number 1, pages 15–28, Feb. 2022.
You can download the Project files here: Download files now. (You must be logged in).





Responses